A brief summary of a simple explanation for ancestral myths of longevity and a potential route to restoring a Methuselahian life span.
Humans and a number of our primate relatives are unusually long-lived compared to most species of mammals. Some people have made it to 120 years or more in recent times, and expecting to reach at least your 80th birthday is not unrealistic.
However, it seems for many that this is not long enough and research into extending longevity has become a major industry. Interestingly, there are myths of extreme longevity implying ancestral life spans that defy reason.
One factor that has emerged as a guide to longevity is the physiological age of the brain. Simplistically, the physiological age of the brain dictates the physiological age of everything else. This is due to the physiological age of the brain being reflected in the degree of juvenility or maturation of the hormonal or endocrine regime it runs.
The degree of juvenility or maturation of the endocrine system is governed by the physiological age of the brain. The degree of juvenility or maturation of the endocrine system, in turn, governs the physiological age of the whole organism including the brain itself. A tight circular mechanism, hence the term 'neuro-endocrine system'. In most species, sexual maturation and reproduction is the end game of biological organisms, a mechanism evolved and regulated the necessary sexual maturation processes by the cycling back of the initially very juvenile fetal neuroendocrine system. Sexual maturation is both the end of the juvenile phase and when normal ageing begins as essentially the primary objective of reproductive capacity has been achieved.
Research efforts to extend longevity by hacking individual bits of our genetic code or modifying our chromosome telomeres etc., may be likened to trying to hold back time by tinkering with the mechanisms of a clock.
It would make more sense to re-transcribe our entire genome in such a way that kept us in a state of near-perpetual juvenility. Strangely enough, this is exactly what happened in our evolutionary past. Our ancestral symbiotic relationship built us an advanced neural time machine where we could exist out of phase with normal ageing for greatly extended periods.
Mythological hints of extreme ancestral longevity are rooted in a very rare and unusual symbiotic relationship that revolved around the ingestion of swollen ovaries that permanently infused our developmental and transcription environment with juvenilising reproductive biochemistry.
Scientific data has provided solid evidence that our distant ancestors were involved in a seed disseminating symbiotic relationship with the angiosperms for tens of millions of years. In addition, we retain an unusual number of juvenile traits that make us look more like a stretched giant-brained fetus than a typical mature mammal. These very unusual aspects of our evolution and physiology are connected by very simple and well understood biological mechanisms that have gone unnoticed due to our current focus on adaptive selection as the primary means of explaining our origins.
During my research into human neural evolution and developing my proposal that our expanded neo-cortex is an emergent structure of symbiotic origin, it appeared to me that a very simple mechanism for our current longevity and potentially much greater ancestral longevity had slipped through our perceptual net.
It may offer a simple route back to greater longevity with much more robust health and advanced perceptual abilities.
When you dig a little deeper you find our 50 million year plus symbiotic relationship was utterly bizarre. It revolved around an increasing specialisation in the ingestion of swollen ovaries. I will say it again, as it can take a while to sink in. For tens of millions of years our ancestors ingested swollen ovaries as part of a co-evolving symbiotic relationship with the flowering plants. The symbiotic relationship was powerful enough to shape and sculpt our basic physiology into that of a frugivorous and still partially arboreal primate. The symbiotic relationship includes very powerful transcription and developmental mechanisms than can easily explain the colossal anomaly that is our neo-cortex and why we may have lived extraordinarily long and healthy lives.
Plant reproductive systems are significantly more juvenilising than mammalian reproductive systems, and being in a symbiotic relationship 'effectively' immersed our mammalian physiology, reproductive and neural system in the reproductive system of the angiosperms for millions of years. This basically kept us in a state of perpetual gestation for the majority of our evolutionary past and facilitated the expansion of our mammalian brain by amplifying and extending the normal neural proliferation window. What emerged wasn't just an expanded version of our mammalian brain, but a completely new kind of brain, a slowly accelerating proliferation of increasingly juvenile and eventually almost embryonic neural tissue.
As a direct result of our utterly weird symbiotic relationship, our ancestors acquired an extremely juvenile, almost embryonic neocortex. It was what is termed an emergent structure, a symbiotic hybrid brain borne of a symbiotic reproductive system. Its structural integrity and function were entirely dependent on the symbiotic relationship being maintained, as would be the case for any emergent symbiotic structure.
Being almost embryonic in its physiology, and as the new outer layer or neo-cortex assumed an executive role, it conferred a raft of rare functional and perceptual traits including slowing the rate of typical maturation and ageing processes to a glacial pace.
The outcome; a direct blanket juvenilising effect of the symbiotic relationship and the additional and increasingly powerful blanket effect of an expanding and increasingly juvenile neo-cortex that resulted in a slowly accelerating extension of longevity by inhibiting the typical maturational mechanisms that result in normal ageing. More simplistically, a juvenilising feedback loop that can explain the accelerating expansion of our neocortex and its tripling in size during the latter stages of the symbiotic relationship.
Again with some pictures; Quite literally a rejuvenating symbiotic relationship.
If our neural system developed in an extremely juvenile reproductive system, it would result in an extremely juvenile brain, the juvenile brain would take much longer to age and keep everything more juvenile including itself for much longer.
Essentially the symbiotic relationship kept us in a state of perpetual gestation in a reproductive system that is more juvenile than our own mammalian reproductive system.
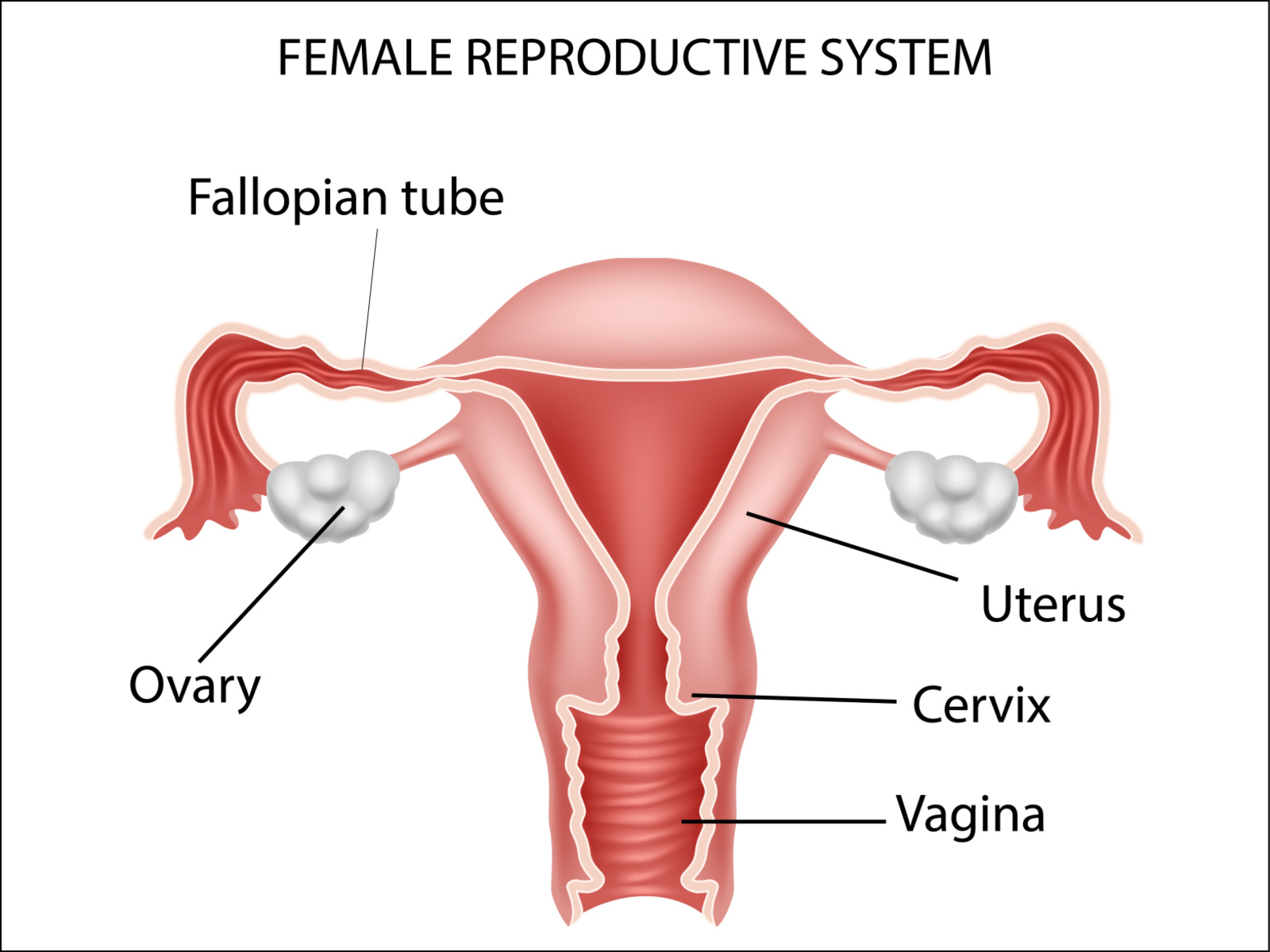 =
= 
Mammalian reproductive systems evolved to provide a slowly maturing transcription and development environment suitable for embryo formation through to a fully developed and functional juvenile.
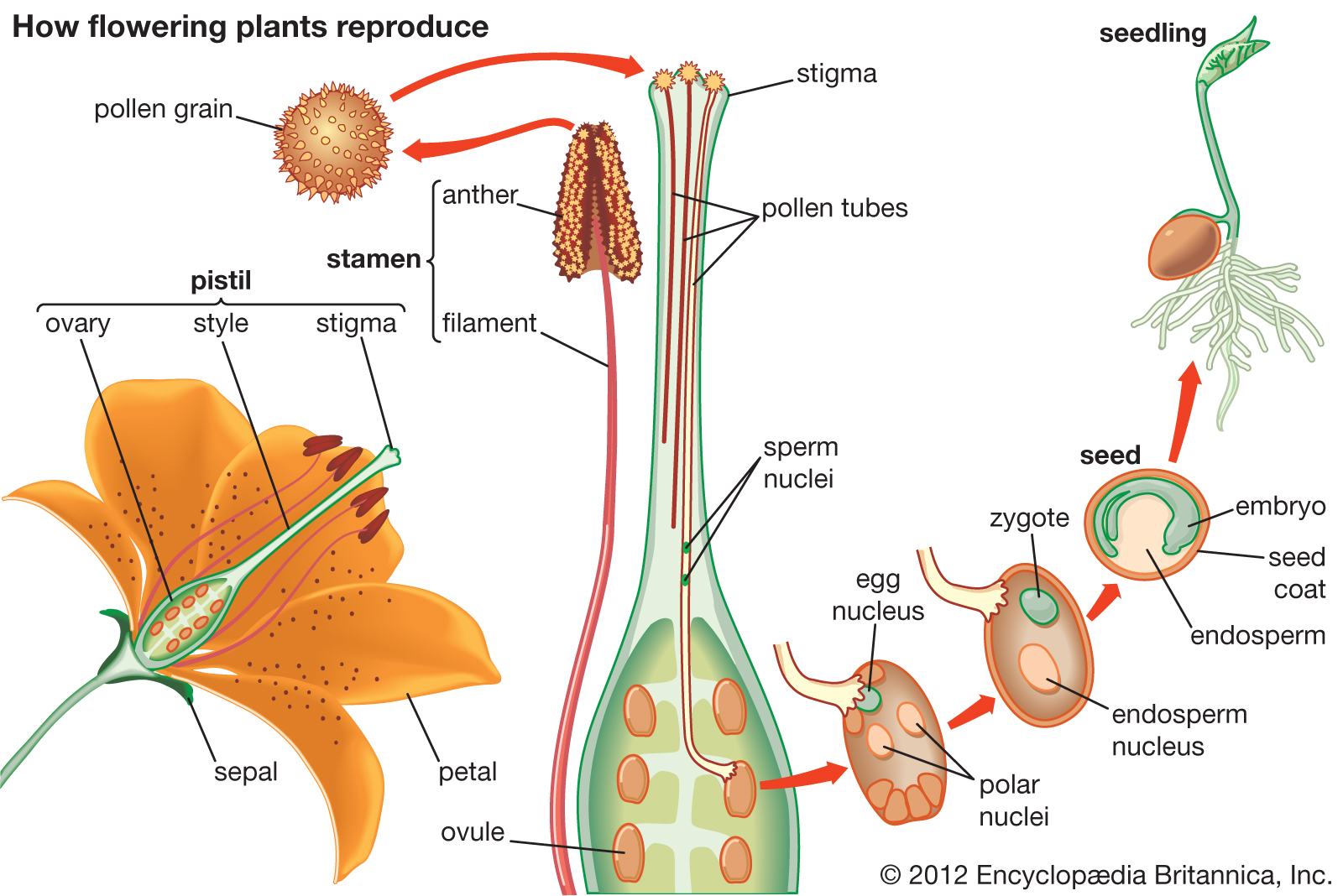 =
= 
Angiosperm reproductive systems evolved to provide a transcription environment suitable for embryo formation and a short period of very limited embryonic development.
Mammalian reproductive systems provide a relatively maturing transcription and developmental environment.
Angiosperm reproductive systems provide a relatively juvenilising transcription and developmental environment.
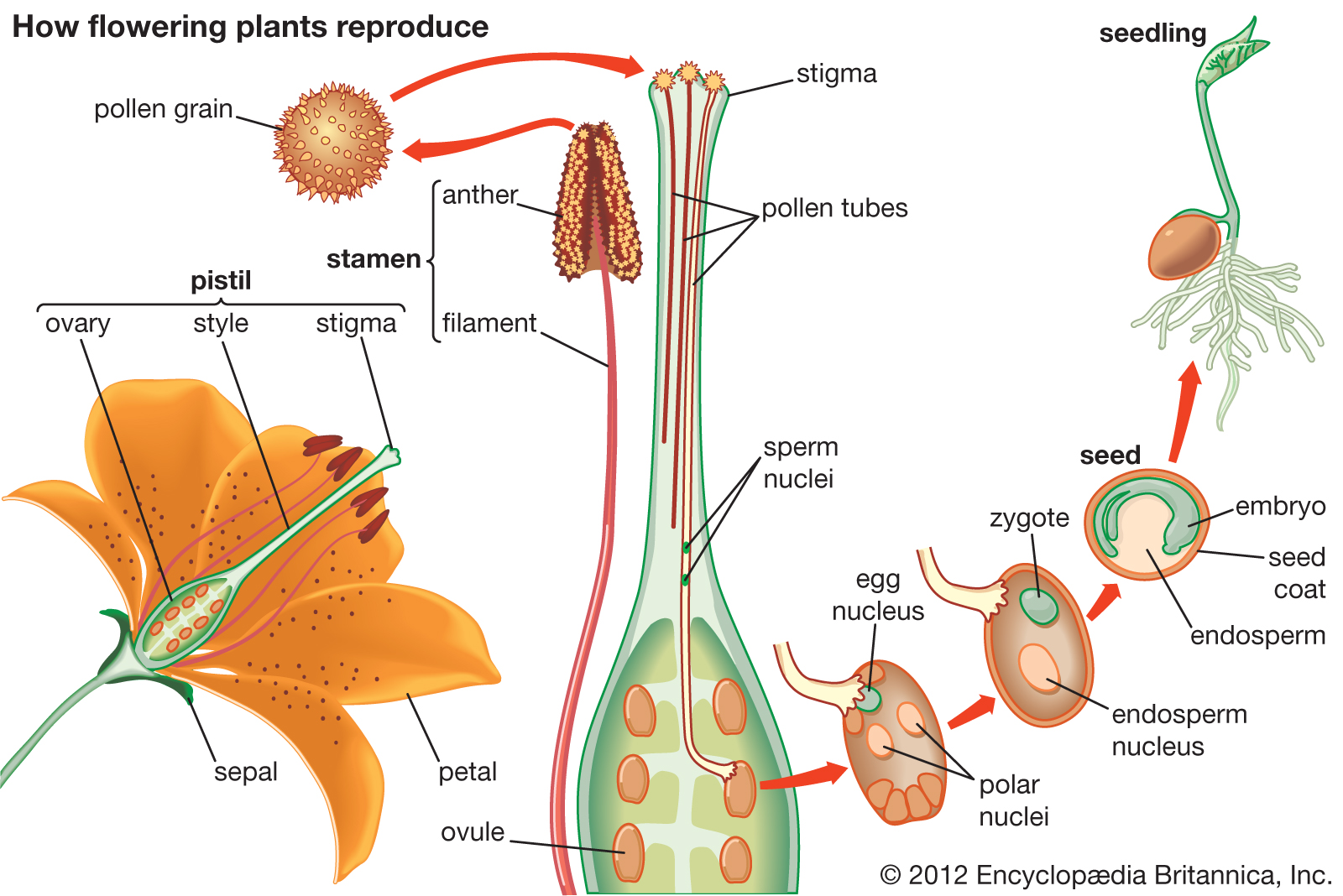
{kind=link}
Two reproductive systems.
So according to the accepted data, our neural system co-evolved and developed in a hybrid reproductive system, part mammal, part plant.
For tens of millions of years our ancestors were in a seed disseminating symbiotic relationship with the reproductive system of the angiosperms via the ingestion of their swollen ovaries.
The physical reproductive system was mammalian, but it was perpetually infused by the more complex and juvenilising reproductive biochemistry of the plant reproductive system.
+  =
= 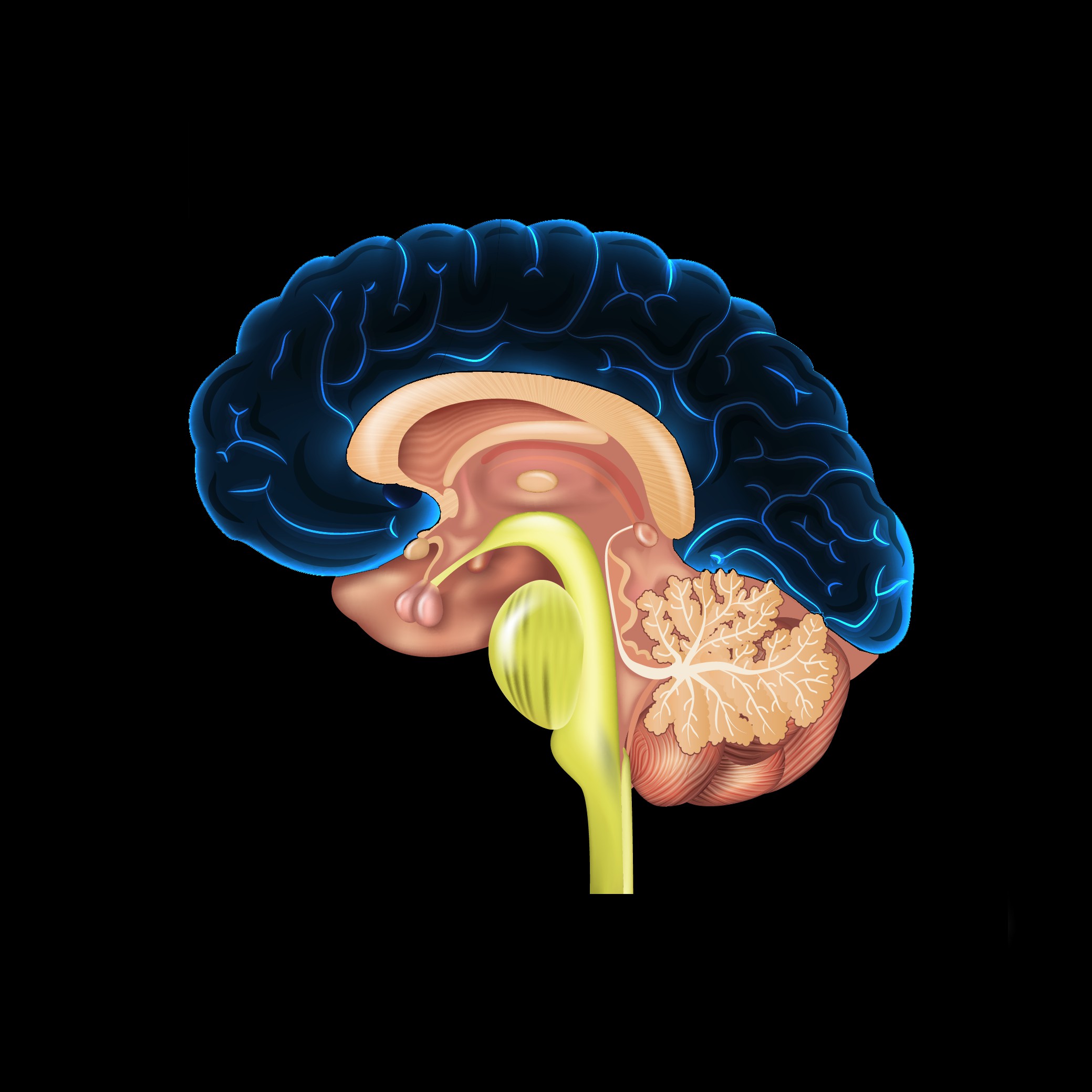
This rare state of perpetual gestation facilitated the proliferation of increasingly juvenile and eventually almost embryonic neural tissue. The juvenilising effect of the angiosperm reproductive system combined with the juvenilising effect of the emerging neo-cortex slowed all stages of typical mammalian maturation, differentiation and ultimately ageing. As long as the relationship was stable, an incrementally minuscule exponential juvenilising feedback loop could eventually accelerate into a runaway feedback loop. This results in a uniquely rapid and accelerating neural expansion of increasingly juvenile and near embryonic neural tissue that in time becomes the most powerful juvenilising factor.
- = 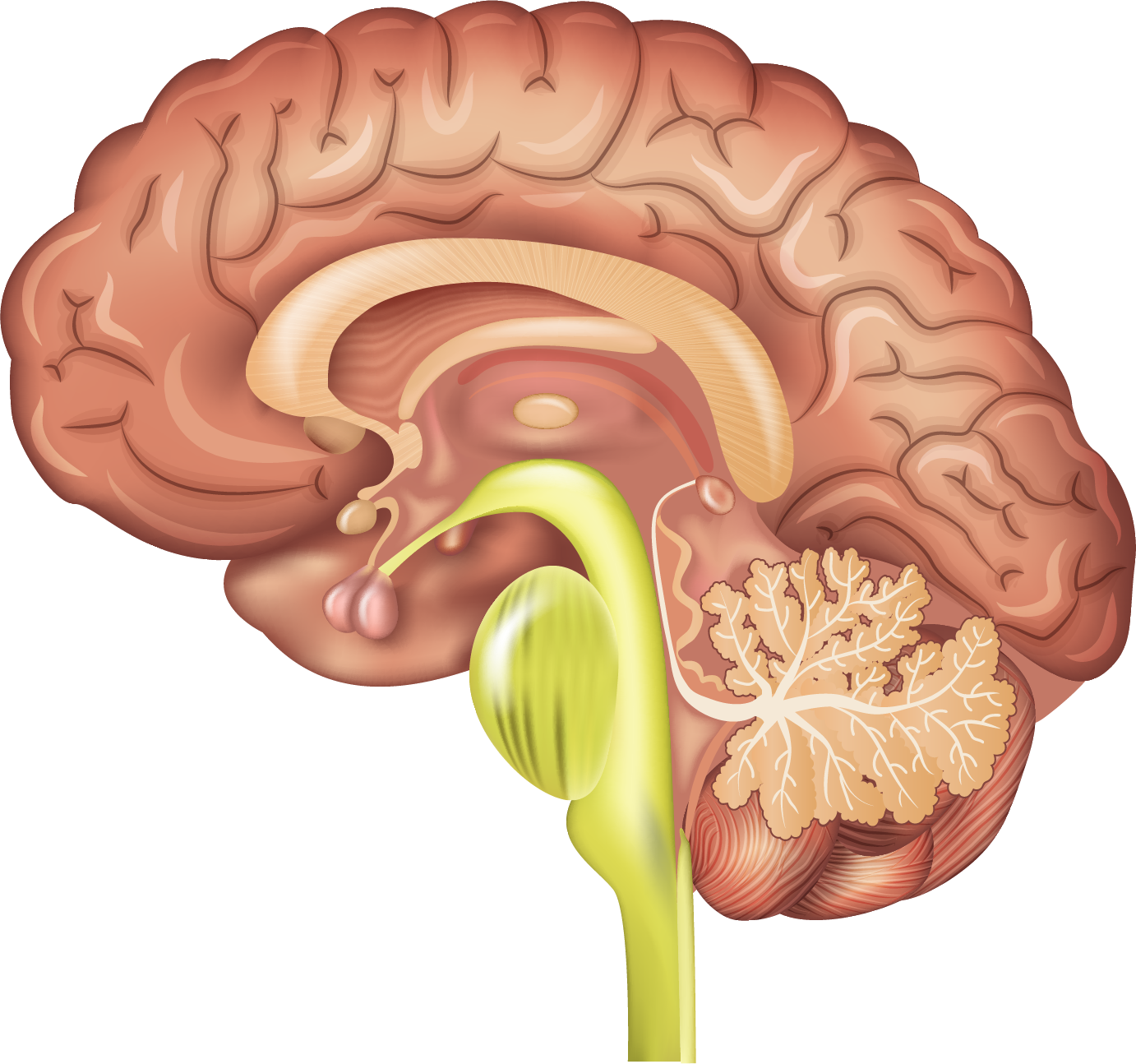
Any breakdown of the symbiotic relationship results in the extremely juvenile neural tissue being exposed to the relatively ageing transcription environment of the stand-alone mammalian reproductive system. The neural embryonic tissue begins to mature and differentiate; essentially it begins ageing even in the uterus without the juvenilising effect of the angiosperm reproductive system. As the new brain ages it accelerates its own ageing, an accelerating maturational feedback loop is initiated, the opposite of the juvenilising feedback loop the symbiotic relationship initiated.
Relative to its symbiotic origins the embryonic new brain differentiates and is structurally transformed, slowly becoming more like the typical mammalian brain tissue it emerged from. Perceptually it also starts to suffer a from a form of senile dementia that begins in the uterus and accelerates during what are considered 'normal' maturational windows. This post symbiotic reversion process includes a slow return to more typical mammalian ageing and longevity. While we still retain some degree of atypical mammalian juvenility, we are at the wrong end of the accelerating maturational feedback loop.
Ultimately the emergent embryonic neural system cannot structurally survive the accelerating maturational or ageing feedback loop and begins to atrophy and erode.
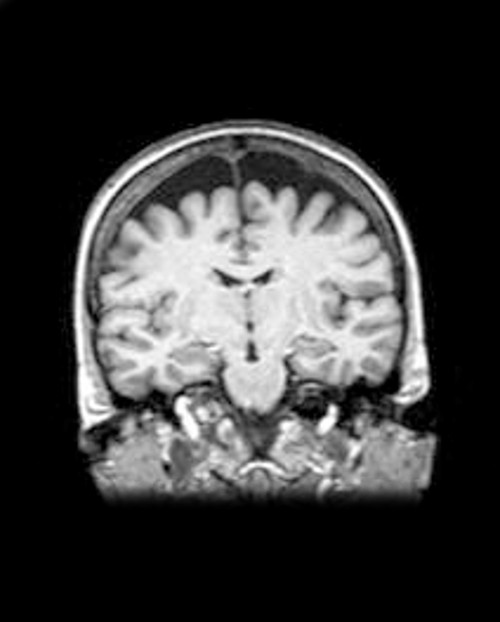
Image linked to paper documenting late-stage atrophication and withering of our new brain.
Attempting to hack longevity while ignoring the catastrophic atrophication of our embryonic new brain is a dementia-like symptom of failure in the advanced perceptual and cognitive capacity it once endowed our Methuselahian ancestors with.
Bit of context.
This may sound a bit unlikely until you sift through the relevant evidence;
For example, pharmacological data from research into the impact of angiosperm reproductive biochemistry on mammalian physiology has classified typically occurring compounds as endocrine disruptors. Specifically, they have a modulating or inhibitory effect on mammalian developmental or 'ageing' hormones such as oestrogens and testosterone. The data supports the general conclusion of a net juvenilising impact on mammalian physiology. Being that these compounds were an integral part of our symbiotic physiology and developmental environment for tens of millions of years, it is hard to avoid the conclusion that they played a significant role in the strange juvenile traits we still exhibit.
More context here.
Too far fetched?
The mechanisms and data exist if you look for them. In fact, it is mostly college-level biology. It simply requires finding a pathway through the existing academic data that nobody bothered looking for.
Interestingly a parallel symbiotic relationship resulting in traits of slowed development and unusual neural expansion appears to suggest a similar mechanism at work in the largest fruit bats.
As with similar traits in frugivorous primates no account is taken of the inevitable hormonal impact on mammalian physiology that would result from ingesting swollen plant ovaries over evolutionary time scales.
Link to pdf Relative Brain Size and Feeding Strategies in the Chiroptera
Genomic analysis of male puberty timing highlights shared genetic basis with hair colour and lifespan
Hot flash: Women who start menstruation and menopause later more likely to live to 90
Intellectual ability and cortical development in children and adolescents
Ageing induced neuro-chemical cerebral asymmetry.
Love to win or hate to lose? Asymmetry of dopamine D2 receptor binding predicts sensitivity to reward vs. punishment
Developmental Changes in Dopamine Neurotransmission in Adolescence: Behavioral Implications and Issues in Assessment
Dopamine-D2-Receptor Blockade Reverses the Association Between Trait Approach Motivation and Frontal Asymmetry in an Approach-Motivation Context
Flavonoids in Treatment of Chronic Kidney Disease
Scientists may have discovered how to slow down aging in humans
Ageing-associated changes in transcriptional elongation influence longevity
Several years ago I ran the basic symbiotic theory past one of the most knowledgeable and sceptical minds I could find in the relevant academic disciplines.
Being that it is in many ways quite a simple theory built on existing mechanisms and data, any significant flaws would be easy to spot by anyone with relevant knowledge.
After several months of pondering, this was the response:
‘This is a totally new way of looking at the evolution of the human brain. It is so totally fresh, unexpected and hitherto un-thought-of that it will probably take a long time before evolutionary anthropologists and psychologists begin to take it on board; but it will make an impact, of that there is no doubt. It will be, it must be, taken very seriously in any discussion of human origins.’ (14/03/2008)
Colin Groves- Professor of Biological Anthropology at the School of Archaeology & Anthropology, Australian National University and author of several books including A Theory Of Human And Primate Evolution.
Please support this project on Patreon @ 'Children of the Forest'; Restoring Our Symbiotic Brain & Mind.
Copyright©2020 Tony Wright